
Tái phân bổ năng lượng giúp tôm thẻ chân trắng Litopenaeus vannamei phản ứng lại stress nhiệt
Tóm tắt
Nhiệt độ đóng vai trò là yếu tố môi trường quan trọng trong hệ sinh thái. Hiểu được sự hợp tác của các mô khác nhau của động vật phản ứng với stress nhiệt là cơ sở để làm rõ cơ chế điều hòa của loài khác nhau khi bị stress nhiệt. Trong đây, phân tích phiên mã so sánh được tiến hành trên ba mô (gan tụy, mang và cơ) của tôm thẻ chân trắng Thái Bình Dương Litopenaeus vannamei khi bị stress nhiệt. Ba các mô biểu hiện các kiểu biểu hiện gen riêng biệt, cho thấy sự hợp tác dựa trên sự phân công có thể đã xảy ra trong số đó. Ở gan tụy và mang, các gen liên quan đến việc tạo ra và sử dụng ATP quy định chặt chẽ và quá trình tạo ra protein tốn nhiều năng lượng gần như đã ngừng hoạt động. Khi ở trong cơ, gen liên quan đến việc tạo và sử dụng ATP, và những thứ liên quan đến một số quy trình tiêu thụ năng lượng đã được điều chỉnh lại. Đồng thời, sự tích lũy đáng kể ATP và giảm tổng nồng độ protein đã được được phát hiện ở gan tụy và mang, trong khi ngược lại ở cơ. Vì vậy, chúng tôi đề nghị rằng các mô khác nhau có thể hợp tác đồng thời với nhau thông qua việc tái phân bổ năng lượng để ứng phó với stress nhiệt. Ít năng lượng hơn chuyển vào quá trình luân chuyển protein ở mang và gan tụy, đồng thời cần nhiều năng lượng hơn cho cơ. Nghiên cứu này không chỉ cung cấp sự hiểu biết toàn diện về cơ chế phân tử của L. vannamei để đáp ứng với nồng độ cao nhiệt độ mà còn đặt nền tảng cho việc khai thác các gen chịu nhiệt và đề xuất các chiến lược hiệu quả để đối phó với môi trường nhiệt độ cao
Giới thiệu
Sự nóng lên toàn cầu ngày càng tăng đã ảnh hưởng đến các sinh vật ở cấp độ sinh thái, sinh lý, tế bào và phân tử (Stock, 2009). Đặc biệt, các sinh vật sống dưới nước, phần lớn là động vật biến nhiệt, dễ bị ảnh hưởng bởi sự thay đổi nhiệt độ hơn (Borgwardt và cộng sự, 2018). Nhiệt độ cao có thể ức chế tốc độ tăng trưởng, giảm số lượng trứng đẻ lần đầu (Sanpradit và cộng sự, 2020), thay đổi cấu trúc cộng đồng (Dziuba và cộng sự, 2020), kéo dài thời gian phát triển (Doan và cộng sự, 2019) và ngoài ra còn làm giảm chức năng miễn dịch của giáp xác (Guo và cộng sự, 2020; Shi và cộng sự, 2020; Zheng và cộng sự, 2018).
Các sinh vật dưới nước thích nghi với nhiệt độ cao thông qua một số chiến lược ở cấp độ tế bào và phân tử, có thể tóm tắt như sau: Tạo ra các protein sốc nhiệt (HSP) hoặc con đường ubiquitin-proteasome tạo điều kiện thuận lợi cho quá trình tái gấp cuộn của các protein bị hư hỏng/biến tính (Wickner và cộng sự, 1999); Sự gia tăng hoạt động của enzyme chống oxy hóa, chẳng hạn như superoxide effutase (SOD), catalase (CAT), glutathione peroxidase (GPX) và glutathione S-transferase (GST), giúp loại bỏ các loại oxy phản ứng (ROS), giải độc các sản phẩm phụ độc hại và điều chỉnh stress oxy hóa (Liu và cộng sự, 2018; Madeira và cộng sự, 2018; Matozzo và cộng sự, 2011; Zheng và cộng sự, 2019); Sự chuyển đổi của quá trình chuyển hóa năng lượng, chẳng hạn như tăng cường chuyển hóa axit béo và tạo đường (Teranishi và Stillman, 2007; Zheng và cộng sự, 2019).
Nhiệt độ đóng vai trò là yếu tố môi trường quan trọng trong hệ sinh thái. Hiểu được sự phối hợp của các mô khác nhau của động vật để ứng phó với stress nhiệt là cơ sở làm rõ cơ chế điều hòa của các loài khác nhau khi bị stress nhiệt.
Trên thực tế, việc điều chỉnh cân bằng năng lượng, đặc biệt là tiêu hao năng lượng và phân bổ năng lượng cho các quá trình gây ra nhiệt nói trên, là nền tảng cho khả năng thích ứng và chịu đựng căng thẳng của sinh vật (Sokolova và cộng sự, 2012).
Khi bị stress nhiệt, các con đường tế bào liên quan đến việc điều chỉnh cung cấp và tiêu thụ năng lượng có trách nhiệm duy trì cân bằng nội môi năng lượng tế bào (Khan và Khan, 2008). Việc điều chỉnh năng lượng trong quá trình thích ứng với nhiệt độ cao tuân theo quy tắc ngân sách năng lượng động (DEB) (Kooijman, 2000).
Ở động vật không xương sống dưới nước, trong điều kiện bình thường (khoảng tối ưu), việc cung cấp ATP thông qua quá trình trao đổi chất hiếu khí có thể trang trải mọi hoạt động, phát triển, tăng trưởng, tái sản xuất và chi phí năng lượng tương ứng một cách đầy đủ.
Trong điều kiện căng thẳng vừa phải (khoảng cho phép) , sự cân bằng năng lượng được phục hồi thông qua sự đánh đổi năng lượng để duy trì tăng lên (ví dụ: protein sốc nhiệt và chất chống oxy hóa) gây tổn hại cho các quá trình khác như tăng trưởng và sinh sản. Trong các tình huống căng thẳng quá mức, quá trình yếm khí một phần được kích hoạt để bù đắp cho việc cung cấp năng lượng hiếu khí không đủ và tốc độ trao đổi chất bị ức chế để đảm bảo cân bằng năng lượng là dừng lại chức năng đòi hỏi ATP (ví dụ: tổng hợp protein) để đảm bảo sự sống còn ngay lập tức. Trong phạm vi gây chết, sự cân bằng giữa cung và cầu ATP bị phá vỡ dẫn đến cái chết của sinh vật (Sokolova và cộng sự, 2012).
Giá trị kinh tế cao và đặc tính sinh sản tuyệt vời giúp tôm thẻ chân trắng Thái Bình Dương Litopenaeus vannamei trở thành một trong những loài nuôi trồng thủy sản chính trên thế giới (FAO, 2020, https://www.fa o.org/publications/sofia/2020/en/ ). Tuy nhiên, vào mùa hè, thời tiết nắng nóng liên tục hoặc nhiệt độ định kỳ trên 35oC dẫn đến tỷ lệ tôm chết cao (Dall và cộng sự, 1990).
Vì vậy, việc nghiên cứu cơ chế phản ứng của L. vannamei với nhiệt độ cao là rất cấp thiết. Hiện tại, hầu hết các nghiên cứu về stress nhiệt chỉ giới hạn ở các mô hoặc chỉ số đơn lẻ, ví dụ, hoạt động của enzyme chống oxy hóa đã bị thay đổi (Estrada-C'ardenas và cộng sự, 2021; Gonz' alez-Ruiz và cộng sự, 2020), HSP mở rộng được tạo ra (T. Chen và cộng sự, 2018; Qian và cộng sự, 2012) và mức tiêu thụ oxy tăng lên đáng kể (Ulaje và cộng sự, 2020). Tuy nhiên, hiểu biết về sự hợp tác của các mô khác nhau và mối quan hệ của nó với năng lượng để ứng phó với stress nhiệt vẫn còn khan hiếm.
Trong tài liệu này, phân tích phiên mã so sánh được tiến hành trên ba mô của tôm bị stress nhiệt, bao gồm gan tụy, mang và cơ. Các gen biểu hiện khác biệt (DEG) của mỗi mô được chiết xuất và đưa vào phân tích làm giàu chức năng. Sau đó, mạng lưới tương tác protein-protein (PPI) được xây dựng để xác định các phân tử chiếm ưu thế. Cho phép những thay đổi lớn của gen khi bị stress nhiệt có liên quan chặt chẽ đến năng lượng, sự biểu hiện của các gen liên quan đến năng lượng, bao gồm tổng hợp ATP, xúc tác ATP, chu trình đường phân và TCA đã được phân tích sâu hơn để khám phá mối quan hệ giữa năng lượng và những thay đổi do nhiệt gây ra. Cuối cùng, tổng nồng độ protein và nồng độ ATP của ba mô đã được phát hiện để xác nhận việc ngừng luân chuyển protein và tái phân bổ năng lượng khi bị stress nhiệt. Những kết quả này cung cấp nguồn thông tin quý giá để phân tích cơ chế thích ứng của tôm để ứng phó với stress nhiệt.
Nghiên cứu này cung cấp những hiểu biết sâu sắc về cơ chế điều tiết của tôm và các động vật biến nhiệt khác để ứng phó với căng thẳng môi trường.
Thảo luận
Trong nghiên cứu này, chúng tôi đã xác định được 746, 1302 và 430 DEG trong gan tụy, mang và cơ của tôm thẻ L. vannamei khi bị stress nhiệt (33 ◦C), tương ứng. Trong khi đó, ít DEG nhưng có chức năng làm giàu tương tự kết quả được chia sẻ giữa ba mô, suy ra rằng phản ứng Cơ chế gây ra stress nhiệt là khác biệt nhưng có tính liên kết ở những mô. Hơn nữa, chúng tôi đã chứng minh sự phân công và lao động của các mô khác nhau của tôm phản ứng với stress nhiệt thông qua so sánh phân tích phiên mã. Nghiên cứu này cung cấp những hiểu biết sâu sắc về quy định cơ chế của tôm và các động vật biến nhiệt khác để đáp ứng với môi trường căng thẳng.
1. Tế bào mang chuyển sang hô hấp kỵ khí và đối phó với Căng thẳng oxy hóa do stress nhiệt
Nhiệt độ cao thường làm tăng tốc độ hấp thụ oxy và tốc độ hô hấp (Ponce-Palafox và cộng sự, 1997; Wang và Chen, 2006), tạo ra ROS quá mức và dẫn đến stress oxy hóa ở động vật giáp xác (Xiong và cộng sự, 2020; Zhai và cộng sự, 2021). Nói chung, động vật thủy sản sử dụng hệ thống chống oxy hóa, chủ yếu bao gồm GSH/GSSG và các enzyme chống oxy hóa, để loại bỏ ROS dư thừa khỏi làm hỏng cấu trúc và chức năng của axit nucleic và protein để thích nghi với môi trường (Welker et al., 2013). Trong khi đó, ROS quá mức cũng dẫn đến phản ứng của hệ thống miễn dịch (Schieber và Chandel, 2014). Căng thẳng nhiệt cấp tính đã được báo cáo là làm giảm hoặc rối loạn khả năng miễn dịch của L. vannamei và giảm khả năng kháng bệnh mầm bệnh trong thời gian ngắn (Cheng và cộng sự, 2005; Wang và cộng sự, 2006). Nó là kết quả của sự ức chế nhiều phân tử miễn dịch trong mang, chẳng hạn như lớp vỏ, ALF, PPAE, prophenoloxidase, lectin loại C, v.v. (Zheng và cộng sự, 2019), những phân tử này cũng được quy định đáng kể trong nghiên cứu này. Sau một thời gian điều hòa miễn dịch, tôm sẽ hình thành khả năng thích ứng miễn dịch (Pan và cộng sự, 2007).
Hơn nữa, nhiệt độ cao không chỉ làm giảm lượng oxy hòa tan trong nước mà còn làm hỏng cấu trúc mang và giảm khả năng trao đổi khí của nó (Xiong và cộng sự, 2020; Zhai và cộng sự, 2021). Do đó, nhiệt độ cao thường đi kèm với tình trạng thiếu oxy (Vaquer-Sunyer và Duarte, 2008). Trong nghiên cứu này, việc kích hoạt con đường HIF-1 (Hình 2A, Bảng S10) tiết lộ rằng rõ ràng mang tôm đã chuyển đổi hô hấp hiếu khí sang hô hấp kỵ khí, điều này được chứng minh rõ hơn bằng sự điều chỉnh tăng của lactate dehydrogenase (một chỉ số về hô hấp kỵ khí) và chu trình TCA bị hạn chế. Một cách nhất quán, chu trình TCA và chuỗi hô hấp bị ức chế, đồng thời tốc độ tạo ATP giảm khi nhiệt độ trong mang của Marsupenaeus japonicus (Zheng và cộng sự, 2019) và Scylla serrata tương ứng tăng lên (Paital và Chainy, 2014). Trong tình huống này, quá trình glycolysis được điều chỉnh tăng cho thấy quá trình chuyển hóa kỵ khí được tăng cường gây ra bởi sự hấp thu oxy tăng lên. Tuy nhiên, những kết quả này không thể giải thích sự tích lũy ATP, đặc biệt khi các biểu hiện của ATP synthase bị điều chỉnh giảm. Khi xem xét việc ngăn chặn các quá trình tiêu tốn năng lượng như sinh tổng hợp glycan, chuyển hóa axit amin và xử lý protein trong mạng lưới nội chất, việc tăng nồng độ ATP có thể là do giảm tiêu hao năng lượng, được chứng minh bằng sự điều chỉnh giảm ATPase.
2. Năng lượng được tiết kiệm nhờ việc ngừng tổng hợp protein ở gan tụy.
Nói chung, stress nhiệt ảnh hưởng đến tổng hợp protein ở động vật không xương sống (Teranishi và Stillman, 2007), đề cập đến quá trình tổng hợp protein và thoái hóa protein (Houlihan, 1991). Trong nghiên cứu này, sự thoái hóa protein bị cản trở, được thể hiện bằng con đường proteosome bị ức chế. Ngoài ra, với vai trò là enzyme phân giải protein, sự điều hòa giảm một số protease/proteinase và trypsin cũng ủng hộ kết luận này. Tuy nhiên, các nghiên cứu về ảnh hưởng của nhiệt độ cao đến sự phân hủy protein đã mang lại kết quả khác nhau. Căng thẳng nhiệt đã thúc đẩy quá trình phân hủy protein để phân hủy có chọn lọc các protein bị sai lệch hoặc bị biến tính ở cua sứ Petrolisthes cinctipes (Teranishi và Stillman, 2007), nhưng làm suy giảm khả năng kích hoạt proteasome 26 S trong tế bào động vật có vú (Kuckelkorn và cộng sự, 2000; Pajonk và cộng sự. , 2005). Nghiên cứu hiện tại hỗ trợ các kết quả sau. Quan trọng hơn, quá trình tổng hợp protein cũng bị ngừng hoạt động thông qua quá trình sinh học ribosome, sinh tổng hợp aminoacyl-tRNA và xử lý protein trong con đường ER sắp xảy ra. Hai con đường đầu tiên tham gia vào quá trình dịch mã từ mRNA sang polypeptide và con đường cuối cùng chịu trách nhiệm xử lý các polypeptide mới được tổng hợp (Merrick và Pavitt, 2018).
Nhiệt độ cao đã được phát hiện là có tác dụng ức chế quá trình tổng hợp protein ở nhiều loài (Spriggs và cộng sự, 2010; Teranishi và Stillman, 2007). Trong nghiên cứu này, việc ngăn chặn quá trình tổng hợp protein đã góp phần làm giảm đáng kể nồng độ protein tổng số. Nó còn làm giảm đáng kể mức tiêu hao năng lượng vì quá trình tổng hợp protein đóng vai trò là quá trình tiêu thụ năng lượng chính trong tế bào (Sokolova và cộng sự, 2012). Mặt khác, việc điều chỉnh giảm ATPase càng khẳng định suy đoán này. Trong trường hợp này, sự tích lũy ATP cho thấy nhu cầu năng lượng thấp đối với tế bào gan tụy, được xác nhận bởi quá trình glycolysis, chu trình TCA và ATP synthase được điều chỉnh xuống. Một cách nhất quán, hoạt động của ATP synthetase giảm và hàm lượng ATP tăng đáng kể trong gan tụy của P. cinctipes (Teranishi và Stillman, 2007) và Penaeus monodon (Jiang et al., 2018) ở nhiệt độ cao. Tuy nhiên, các nghiên cứu trước đây đã suy đoán rằng quá trình tổng hợp ATP ở nhiệt độ cao được tăng tốc, góp phần tích lũy hàm lượng ATP, nhưng việc tiêu hao năng lượng có thể bị bỏ qua.
3. Sự cảm ứng lớn của HSP tiêu thụ năng lượng lớn trong cơ
Sản xuất HSP là một trong những đặc điểm nổi bật nhất ở động vật giáp xác khi bị stress nhiệt (Hong và cộng sự, 2020; Liu và cộng sự, 2018; Zheng và cộng sự, 2019). Đóng vai trò là người đi kèm phân tử điển hình, HSP được sử dụng để bảo vệ protein khỏi sự kết tụ thông qua việc tinh chế các protein bị biến tính hoặc làm suy giảm các protein bị gấp sai do stress nhiệt theo cách phụ thuộc ATP (Kregel, 2002). Trong nghiên cứu hiện tại, người ta đã quan sát thấy sự cảm ứng mạnh mẽ của HSP21, HSP40, HSP60, HSP70 và HSP90 trong cơ, điều này phù hợp với các nghiên cứu trước đây (Qian và cộng sự, 2012; Ulaje và cộng sự, 2020). Xem xét vai trò chủ yếu của HSP trong cơ bắp, quá trình phiên mã và dịch mã đi kèm đã được tăng cường đáng kể cho thế hệ của chúng. Những tác động này dẫn đến sự gia tăng đáng kể nồng độ protein tổng số. Cùng với thực tế là chức năng đi kèm của HSP phụ thuộc vào ATP (Yeong và MacRae, 2011), việc xuất hiện những thay đổi trên sẽ tiêu tốn một lượng lớn năng lượng, được xác nhận bằng việc giảm ATP. Kết hợp với chu trình TCA được điều chỉnh tăng, ATP synthase và ATPase, những kết quả này chỉ ra rằng các tế bào cơ bắp đang ở trạng thái nhu cầu năng lượng cao. Tương tự, nồng độ ATP và điện tích năng lượng adenylate (AEC) đều tăng trong cơ cua bơi Portunus trituberculatus dưới nhiệt độ cao (Lu và cộng sự, 2015), chứng minh rằng nhu cầu năng lượng cao của cơ là rất cần thiết cho loài giáp xác để ứng phó với stress nhiệt.
4. Nhiều năng lượng hơn được phân bổ lại để bảo vệ nhiệt giữa ba mô
Trong điều kiện môi trường tối ưu, nguồn cung cấp năng lượng đủ để trang trải mọi chi phí duy trì cơ thể và ngoài ra nó còn cho phép tăng trưởng, sinh sản và tích lũy dự trữ năng lượng. Tuy nhiên, việc duy trì được ưu tiên để bảo vệ căng thẳng và đảm bảo sự sống sót, trong khi dòng năng lượng được định tuyến lại dẫn đến giảm tổng thể các nguyên nhân gây ra hiệu quả sử dụng năng lượng, như ngừng tăng trưởng, tạm dừng sinh sản và huy động các kho dự trữ năng lượng trong môi trường căng thẳng (Klepsatel và cộng sự, 2016). ; Storey và cộng sự, 2011). Nói chung, tôm được lấy mẫu trong nghiên cứu này nằm trong khoảng cho phép và khoảng gây chết dựa trên quy tắc DEB. Các protein sốc nhiệt và chất chống oxy hóa tạo ra rất nhiều trong giai đoạn trước, quá trình yếm khí một phần được kích hoạt và nhiều chức năng đòi hỏi ATP (ví dụ như tổng hợp protein) bị ngừng hoạt động để đảm bảo cân bằng năng lượng cho sự sống sót ngay lập tức trong giai đoạn sau (Kooijman, 2000).
Tuy nhiên, trạng thái nhu cầu năng lượng của ba mô là khác nhau khi bị stress nhiệt. Cụ thể, mang và gan tụy ở trạng thái nhu cầu năng lượng thấp, trong đó cả việc tạo ra và sử dụng năng lượng đều ở mức thấp. Tuy nhiên, tình hình hoàn toàn ngược lại ở cơ khi cả quá trình tạo và sử dụng năng lượng đều được đẩy nhanh, cho thấy cơ đang ở trạng thái nhu cầu năng lượng cao. Thật hợp lý khi cho rằng hiện tượng này là do sự phân bổ lại dòng năng lượng giữa các mô khác nhau. Nghĩa là, ít năng lượng hơn được chuyển vào quá trình tạo ra protein tốn nhiều năng lượng ở mang và gan tụy để duy trì sự sống ở mức tối thiểu, nhiều năng lượng hơn được phân bổ lại để bảo vệ nhiệt trong cơ và hệ thống chống oxy hóa ở mang trong điều kiện nhiệt độ cao (Hình 6)
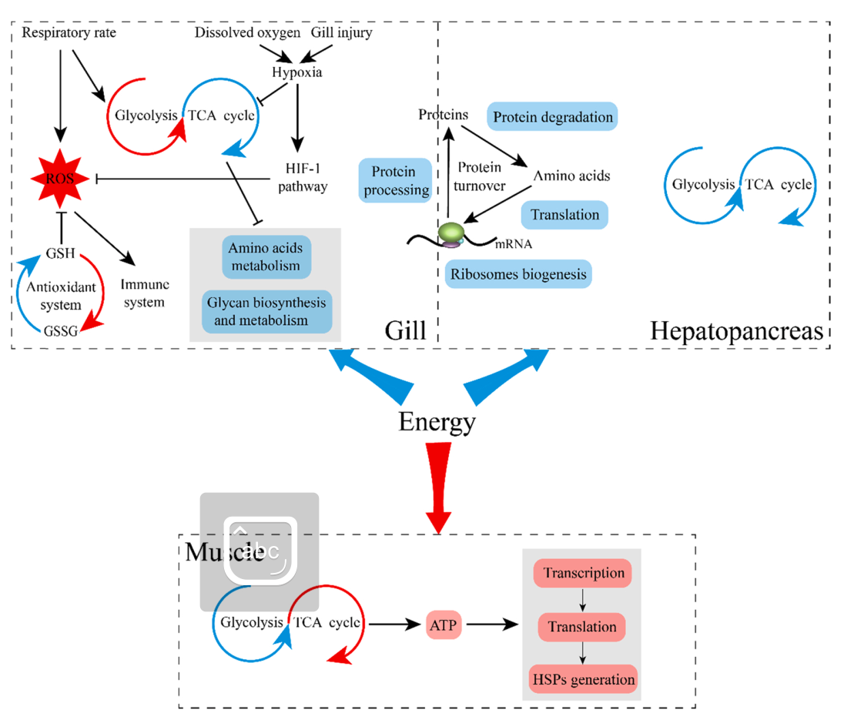
Khác biệt đáng chú ý là việc tạo ra HSP chỉ được quan sát thấy ở cơ thay vì ở gan tụy và mang trong nghiên cứu này. Điều này có thể được giải thích là do HSP đóng vai trò quan trọng trong việc bảo vệ các protein cấu thành cơ, như Actin, myosin và tropomyosin, khỏi sự kết tụ do nhiệt (Markov và cộng sự, 2008). Nhưng làm thế nào ba mô này phối hợp với nhau vẫn chưa được biết, có thể hệ thần kinh đóng vai trò quan trọng trong quá trình điều hòa tích hợp này.
Lược dịch: BP kỹ thuật công ty VPAS
Xiaoxi Zhang a,b, Jianbo Yuan a,b, Xiaojun Zhang a,b,*, Yang Yu a,b, Fuhua Li ; Comparative transcriptomic analysis unveils a network of energy reallocation in Litopenaeus vannamei responsive to heat-stress; Ecotoxicology and Environmental Safety 238 (2022) 113600
- Robins McIntosh - Liệu khủng hoảng tôm có thể đảo ngược được không?
- Oceanloop mở rộng nuôi tôm trong nhà
- Robins McIntosh - Sự gia tăng bệnh tôm
- Phát hiện virus đốm trắng trong 24 giờ
- Thị trường cá rô phi toàn cầu
- Những phát hiện gần đây về nhiễm ký sinh trùng EHP
- Khảo sát cho thấy sự phổ biến của độc tố nấm mốc trong thức ăn chăn nuôi và thủy sản
- Vật liệu sinh học mới có thể giúp chống lại tình trạng kháng kháng sinh ở cá





