
Vibrio parahaemolyticus gây chết tôm
PHẦN I: GIỚI THIỆU
Từ cuối năm 2019 đầu năm 2020 xuất hiện dịch bệnh mới trong ngành nuôi tôm được gọi là bệnh mờ đục hậu ấu trùng (TPD) hoặc bệnh thủy tinh trên tôm giống ở các tỉnh quen biển phía Nam Trung Quốc. Bệnh mờ đục hậu ấu trùng ngày càng trở nên phổ biến trên tôm, gây tổn thất hơn 70% trại ương tôm ven biển ở Trung Quốc vào mùa xuân năm 2020. Vibrio parahaemolyticus có độc lực cao (Vp-JS20200428004-2) là mầm bệnh gây ra TPD và được gọi là V. parahaemolyticus gây bệnh TPD hoặc VpTPD.
VpTPD có khả năng gây chết cao, đặc biệt đối với hậu ấu trùng lúc 4–7 ngày tuổi (PL4–PL7). Tỷ lệ chết tích lũy của hậu ấu trùng bị nhiễm bệnh có thể lên tới 100% trong 3 ngày. Hậu ấu trùng tôm bị nhiễm bệnh có các triệu chứng lâm sàng điển hình như gan tụy nhợt nhạt hoặc không màu, đường tiêu hóa trống rỗng, tôm bệnh trở nên trong suốt (mờ). Do đó, nông dân địa phương đặt tên là “ bệnh mờ đục trên hậu ấu trùng tôm thẻ” hoặc “tôm thủy tinh”. Nhiễm VpTPD ở giai đoạn hậu ấu trùng, tôm (Penaeus vannamei) có thể gây ra những thay đổi mô bệnh học tương tự với với bệnh hoại tử gan tụy cấp tính (AHPND). Các tế bào biểu mô của ống gan tụy và ruột giữa bị hoại tử và bong tróc. Có thể quan sát được một số lượng lớn vi khuẩn cư trú ở gan tụy và ruột giữa dưới kính hiển vi. Trong khi đó, độc tính của VpTPD (vp-HL-202005) đối với hậu ấu trùng P. vannamei cao hơn khoảng 1.000 lần so với chủng V. parahaemolyticus gây AHPND.
Cho đến năm 2023, tỷ lệ mắc bệnh TPD vẫn còn phổ biến ở các trại ương và trang trại nuôi P. vannamei ở các tỉnh ven biển Trung Quốc. Mặc dù một số loại kháng sinh được báo cáo là có thể tiêu diệt hoặc ức chế VpTPD, nhưng nhu cầu sản xuất tôm không có kháng sinh, các biện pháp an toàn sinh học được ưu tiên, bao gồm phát hiện sớm và xử lý khử trùng để ngăn chặn sự xuất hiện và phát triển của TPD. Do đó, nhu cầu cấp thiết là phải điều tra yếu tố độc lực chính của VpTPD để phát triển các kỹ thuật chẩn đoán hiệu quả và các chiến lược phòng ngừa TPD tiếp theo.
Trong nghiên cứu hiện tại, chúng tôi tiến hành nghiên cứu các protein độc lực quan trọng có trọng lượng phân tử khác nhau góp phần gây bệnh cho VpTPD đối với hậu ấu trùng thông qua các thử nghiệm cảm nhiễm. Sau đó, các đoạn protein có độc lực là yếu tố độc lực tiềm ẩn của VpTPD được xác định khối lượng bằng phép đo phổ khối và giải trình tự bộ gen. Bên cạnh đó, chúng tôi cũng nghiên cứu sự hiện diện của yếu tố protein có độc lực cao (VHVP) của Vibrio ở các chủng Vibrio phân lập và sự xuất hiện của VHVP ở các trang trại nuôi tôm bị bệnh TPD từ các khu vực địa lý khác nhau của Trung Quốc. Kết quả nghiên cứu làm rõ cơ chế gây bệnh của VpTPD ở giai đoạn hậu ấu trùng của tôm P. vannamei.
PHẦN II: NGUYÊN LIỆU VÀ PHƯƠNG PHÁP
PHẦN III. KẾT QUẢ (tóm lược)
3.1 Khả năng gây bệnh của các yếu tố độc lực của VpTPD được xác định bằng thử nghiệm cảm nhiễm
Tỷ lệ tử vong tích lũy của hậu ấu trùng P. vannamei bị cảm nhiễm với cả VpTPD sống và các phần protein của nó có trọng lượng phân tử khác nhau được thể hiện trong Hình 1b. Trong thời gian thử nghiệm 40 giờ với các nghiệm thức sau:
- Nhóm không cảm nhiễm (đối chứng âm): không ghi nhận trường hợp tôm chết.
- Nhóm cảm nhiễm VpTPD (7,1x105 CFU/mL): Xuất hiện tôm chết sau 8 giờ và tỷ lệ chết đạt 100% sau 24 giờ.
- Nhóm cảm nhiễm VpTPD +U (>100 kDa): Xuất hiện tôm chết lúc 16 giờ và tỷ lệ đạt 90% sau 32 giờ.
- Nhóm cảm nhiễm VpTPD + U (50–100 kDa), VpTPD + U (30–50 kDa), VpTPD + U (10–30 kDa) và VpTPD + U (100 kDa): Tỷ lệ chết không quá 10% sau 32 giờ cảm nhiễm.
- Chỉ có nhóm cảm nhiễm VpTPD+U (>100 kDa) cho kết quả độc lực (khả năng gây chết) tương đồng với VpTPD. Các yếu tố độc lực hiệu quả của VpTPD phải nằm trong phần (MW >100 kDa) của chiết xuất protein bằng phương pháp phá vỡ siêu âm VpTPD + U.
Thí nghiệm này đã chứng minh rằng VpTPD và nhóm protein có trọng lượng phân tử > 100 kDa có khả năng gây bệnh TPD.
3.2. Phân tích mô bệnh học của tôm giai đoạn hậu ấu trùng từ các nhóm cảm nhiễm
Kiểm tra mô bệnh học cho thấy hoại tử nghiêm trọng và bong tróc các tế bào biểu mô ở cả ống gan tụy và ruột giữa của hậu ấu trùng bị nhiễm VpTPD sống sau 24 giờ sau thử thách (Hình 1c). Trong nhóm được cảm nhiễm với VpTPD + U (>100 kDa), quan sát thấy hoại tử nhẹ và bong tróc các tế bào biểu mô ở cả ống gan tụy và ruột giữa ở thời điểm 16 và 24 giờ sau cảm nhiễm, đồng thời xảy ra hoại tử và bong tróc nghiêm trọng các tế bào biểu mô ở thời điểm 32 và 40 giờ cảm nhiễm (Hình 1d). Những thay đổi mô bệnh học nghiêm trọng nhất được thấy ở ruột giữa lúc 32 giờ sau thử thách, và sự hoại tử nghiêm trọng của các tế bào biểu mô làm cho các tế bào biểu mô rơi ra khỏi màng đáy của ruột giữa và phân tán vào khoang của ruột giữa (Hình 1d). Ngược lại, không có thay đổi mô bệnh học rõ ràng ở ống gan tụy và ruột giữa của các cá thể hậu ấu trùng từ nhóm đối chứng (Hình 1c).
Thí nghiệm này đã chứng minh rằng VpTPD và nhóm protein có trọng lượng phân tử > 100 kDa có khả ảnh hưởng đến tế bào biểu mô ở ống gan tụy và ruột giữa của tôm
3.3. Xác định các yếu tố độc lực của VpTPD bằng SDS-PAGE và phân tích khối phổ
Thí nghiệm này đã chứng minh rằng trong nhóm protein có trọng lượng phân tử > 100 kDa, có 3 protein khác nhau: VpTPD_4-2-1, VpTPD_4-2-2 và VpTPD_4-2-3.
- VpTPD_4-2-1 là phức hợp protein độc tố diệt côn trùng
- VpTPD_4-2-2 là protein độc lực
- VpTPD_4-2-3 là aconit hydratase B (không sử dụng cho thí nghiệm tiếp theo vì theo những nghiên cứu trước aconitate hydratase B không phải là protein gây độc)
3.4. Giải trình tự, phân tích và so sánh bộ gen của chủng VpTPD và không VpTPD
Bộ gen hoàn chỉnh của VpTPD bao gồm hai nhiễm sắc thể dạng vòng (Hình 3a) và ba plasmid (Hình 3c). Hai nhiễm sắc thể dạng vòng có chiều dài lần lượt là 3.527.627 bp (nhiễm sắc thể 1) và 1.887.516 bp (nhiễm sắc thể 2). Ba plasmid của VpTPD lần lượt có kích thước là 212.543 bp, 187.791 bp và 60.506 bp. Trong khi những chủng Vibrio parahaemolyticus không gây bệnh TPD (Vp1616) chỉ có 2 nhiễm sắc thể dạng vòng và không có plasmid, 2 nhiễm sắc thể tròn lần lược có kích thước là 3.288.162 bp (nhiễm sắc thể 1) và 1.923.178 bp (nhiễm sắc thể 2). Phân tích so sánh thông tin bộ gen giữa VpTPD và Vp1616 đã chứng minh rằng hai gen yếu tố độc lực giả định (GE005140 và GE005139) chỉ có trong VpTPD chứ không có trong Vp1616. Dựa vào kết quả căn chỉnh nhiều trình tự bằng công cụ Blastx từ NCBI, hai gen yếu tố độc lực giả định (GE005140 và GE005139), được dự đoán mã hóa các yếu tố độc lực I và II, có độ tương đồng lần lượt là 100% và 99,49% với trình tự amino acid của 2 protein độc tố diệt côn trùng (GenBank: WP_269169668.1) và protein độc lực (GenBank: APX09935.1).
3.5. Đặc điểm trình tự của các yếu tố độc lực duy nhất của VpTPD
Dựa trên phân tích khối phổ và bộ gen so sánh, hai protein độc lực giả định là VpTPD_4-2-1 và VpTPD_4-2-2, được mã hóa lần lượt bởi GE005140 và GE005139 trong VpTPD, được đặt tên giả định là VHVP-1 và VHVP-2. Đặc biệt, hai gen yếu tố độc lực (GE005140 và GE005139) chỉ có trong VpTPD chứ không có trong Vp1616. Các gen GE005140 (vhvp-1) và GE005139 (vhvp-2) được tìm thấy nằm song song trên plasmid 187.791- bp của bộ gen VpTPD và được dự đoán là phụ thuộc vào cùng một vùng khởi đầu (promoter)
Theo phân tích công cụ tìm khung đọc mở (ORF) trong số các gen vhvp, VHVP-1 bao gồm 2.544 amino acid, với khối lượng phân tử dự đoán là 283,37 kDa và pI dự đoán là 4,69, và VHVP-2 bao gồm 1.421 gốc amino acid, với khối lượng phân tử dự đoán là 161,34 kDa và pI dự đoán là 4,63.
3.7. Phát hiện VpTPD bằng PCR
Để phát triển phương pháp phát hiện PCR đối với VpTPD, các mồi PCR được thiết kế để nhắm mục tiêu các gen vhvp-1 và vhvp-2 (Hình 4a; Bảng 3). Cả hai mẫu DNA từ mẫu phân lập VpTPD và mô tôm bị TPD đều có thể được khuếch đại và sản phẩm có kích thước 362, 351 và 303 bp bằng cách sử dụng VpTPD-vhvp-1-F1/R1, VpTPD-vhvp-2-F1/R1 và VpTPD -vhvp-2-F2/R2 bộ mồi tương ứng (Hình 4a). Phân tích độ đặc hiệu của các mồi được thực hiện bằng cách sử dụng các mẫu DNA từ các chủng không phải VpTPD, bao gồm V. parahaemolyticus-0421B, Pseudoalteromonas flavipulchra (CDM8), V. parahaemolyticus gây AHPND (VpAHPND, 20200610006-16), V. alginolyticus (20150606001- 2), V. harveyi (20170902102-3), V. owensii (20150709001-2) và V. campbellii (20150606027-2). Kết quả cho thấy không có sản phẩm PCR nào được khuếch đại khi sử dụng DNA từ các chủng không phải VpTPD làm mẫu, điều này cho thấy bộ mồi PCR chỉ mang tính đặc hiệu cho VpTPD (Hình 4b).
Bảng 3: Cặp mồi PCR phát hiện gen vhvp để nhận diện VpTPD

3.8. Phân tích dịch tễ học của VpTPD
Tổng số 179 mẫu tôm được thu thập từ các trang trại nuôi tôm khác nhau ở Trung Quốc. Các cuộc điều tra dịch tễ học thực địa và phân tích mô bệnh học trong phòng thí nghiệm cho thấy TPD xảy ra ở các trang trại nuôi tôm ở các tỉnh Hà Bắc, Sơn Đông, Giang Tô, Hải Nam và Tân Cương (Hình 4c). Tất cả các mẫu DNA được ly trích từ 179 mẫu tôm đều được điều tra dịch tễ học phân tử bằng cách sử dụng phương pháp PCR xét nghiệm VpTPD. Kết quả PCR cho thấy gen vhvp-1 mục tiêu (chứa các miền VRP1, neurami-nidase và TcA) và gen vhvp-2 (chứa các miền SpvB và TcdB) chỉ có thể được khuếch đại trong các mẫu tôm mắc bệnh TPD chứ không phải từ mẫu tôm khỏe mạnh hoặc không có TPD (Hình 4c). Ngoài ra, chủng V. parahaemolyticus 20211213002-3 được phân lập từ trang trại nuôi tôm ở tỉnh Hồ Nam và chỉ mang gen vhvp-1 nhưng không có gen vhvp-2 và chúng không thể gây tử vong cho hậu ấu trùng P. vannamei thí nghiệm trong cảm nhiễm (Hình 5a). Kết quả cho thấy gen vhvp-2 là gen độc lực quan trọng trong VpTPD.
PHẦN IV: THẢO LUẬN
TPD, một loại bệnh mới nổi chủ yếu ảnh hưởng đến hậu ấu trùng tôm với các hội chứng điển hình là gan tụy và đường tiêu hóa nhợt nhạt hoặc không màu, đã trở thành mối đe dọa cấp bách đối với ngành nuôi tôm ở Trung Quốc (4). Trong một nghiên cứu gần đây đã báo cáo một loài V. parahaemolyticus (VpTPD) mới đã được xác nhận là tác nhân gây bệnh TPD mới nổi dựa trên việc phân lập, xác định và xét nghiệm tác nhân gây bệnh, theo bốn tiêu chí của định đề Koch (4). Tuy nhiên, cơ chế gây bệnh của VpTPD vẫn chưa được hiểu đầy đủ, điều này đã hạn chế việc phòng ngừa và kiểm soát VpTPD trong quá trìnhvnuôi tôm thực tế. Trong nghiên cứu này, chúng tôi đã thực hiện các nghiên cứu chuyên sâu, bao gồm các thử nghiệm thử thách ngâm, phân tích khối phổ, phân tích mô bệnh học và phân tích gen để xác định các yếu tố độc lực cụ thể của VpTPD gây ra bệnh hậu ấu trùng mờ ở P. vannamei. Kết quả cho thấy protein độc tố mới, được chỉ định là VHVP-2 (MW >100 kDa), có chứa các miền được bảo tồn của protein plasmid độc lực Salmonella, protein phức hợp độc tố diệt côn trùng và neuraminidase, là yếu tố độc lực chính của VpTPD (Hình 5c).
Các thử nghiệm cảm nhiễm ngâm trong nghiên cứu hiện tại cho thấy rằng một phần protein cụ thể, trong đó MW >100 kDa từ dịch ly giải của VpTPD, có thể gây chết tôm hậu ấu trùng tương tự như nuôi cấy VpTPD thuần khiết sống. Kết quả này ban đầu chỉ ra rằng yếu tố độc lực của VpTPD phải nằm ở phần protein có MW >100 kDa.Ngoài ra, chất nổi trên bề mặt của môi trường nuôi cấy VpTPD không cho thấy bất kỳ tác động độc lực đáng kể nào đối với hậu ấu trùng tôm so với PBS trong các thử nghiệm thử thách của chúng tôi, điều này chỉ ra rằng protein độc lực chính của VpTPD có thể không được bài tiết trong điều kiện nuôi cấy của nghiên cứu này.
Các nghiên cứu trước đây đã báo cáo rằng SDS-PAGE và phân tích khối phổ đã được áp dụng rộng rãi để xác định các protein độc lực của vi khuẩn với các kích cỡ khác nhau. Trong số ba đoạn protein chính (MW >100 kDa) trong VpTPD dựa trên phân tích SDS-PAGE, hai trong số chúng (VpTPD_4-2-1 và VpTPD_4-2-2) được phát hiện có sự tương đồng trình tự cao với yếu tố độc lực đã biết theo khối lượng phân tích quang phổ và chúng được xác định là yếu tố độc lực tiềm ẩn của VpTPD. Điều thú vị là các protein có tính tương đồng cao của protein độc lực ứng cử viên của VpTPD, bao gồm WP_269169668.1 và APX09935.1, đã được các nhà nghiên cứu khác gửi tới NCBI GenBank vào năm 2017, cho thấy rằng các chủng mang chúng lẽ ra đã bắt đầu lây lan ở một số khu vực trên thế giới trước năm 2017 hoặc sớm hơn. Đối với các chủng vi khuẩn mang protein có độc lực tương đồng, sự phân bố, phương thức lây truyền và tác động gây bệnh của chúng đối với động vật thủy sản rất đáng được nghiên cứu thêm.
Nghiên cứu hiện tại đã chứng minh rằng plasmid chứa gen vhvp độc lực của VpTPD cũng mang traG, traE, traB, traC và các gen liên quan đến chuyển giao liên kết khác. Vì vậy gen độc lực vhvp trong VpTPD có thể truyền qua sự tiếp hợp giữa các loài Vibrio khác nhau (bao gồm V. natriformis, V. Campbellii và V. alginolyticus, được phân lập từ tôm bị bệnh). Nghiên cứu đã phát hiện rằng TPD được gây ra bởi các mầm bệnh khác nhau mang cùng một gen vhvp (gen có thể chuyển khác loài) do đó chúng ta cần chú ý hơn đến VpTPD vì nguy cơ lây truyền ngang cao hơn.
Hơn nữa, các thí nghiệm xóa và bổ sung các đột biến của gen vhvp-2 trong VpTPD đã xác nhận thêm rằng gen vhvp-2 đóng vai trò chính trong việc hiện thực hóa độc lực của VpTPD. Trong khi đó, kết quả điều tra dịch tễ học và thử nghiệm cảm nhiễm cho thấy chủng V. parahaemolyticus phân lập chỉ mang gen vhvp-1 và thiếu gen vhvp-2 không thể gây chết ấu trùng P. vannamei thí nghiệm. Tất cả các kết quả nêu trên chỉ ra rằng vhvp-2 là gen độc lực chính của VpTPD ở P. vannamei. Cơ chế chức năng của yếu tố độc lực VHVP-2 trong việc gây ra sự bong tróc các tế bào biểu mô ruột của tôm nhiễm VpTPD cần được nghiên cứu thêm.
Nghiên cứu hiện tại cho thấy VpTPD trở nên có độc lực gây chết đối với hậu ấu trùng tôm vì nó thu được gen vhvp-2 mã hóa miền độc lực của Salmonella plasmid 28,1 kDa A protein và 65 kDa B protein (SpvB). Trong khi đó, những kết quả này cho thấy tôm bị nhiễm VpTPD có thể gây ra rủi ro tiềm ẩn cho sức khỏe cộng đồng cũng như các động vật nuôi hoặc động vật hoang dã khác thông qua việc lây lan gen vhvp-2 độc hại trong môi trường nước.
Tóm lại, chúng tôi đã chứng minh sơ bộ rằng một loại protein độc lực mới, VHVP-2, là độc tố chính của VpTPD và nó được mã hóa bởi gen vhvp-2 nằm trên plasmid 187.892-bp của bộ gen VpTPD. Điều này có nghĩa là mầm bệnh cơ hội V. parahaemolyticus trở nên độc hại gây chết đối với hậu ấu trùng tôm bằng cách có được yếu tố độc lực của VHVP-2. Ngoài ra, nghiên cứu này đã thiết lập phương pháp phát hiện PCR VpTPD để cảnh báo sớm TPD. Những kết quả này đã chứng minh những hiểu biết mới về cơ chế gây bệnh của VpTPD và cung cấp phương pháp phát hiện phân tử đầu tiên cho VpTPD. Nghiên cứu hiện tại sẽ hữu ích cho việc điều tra sâu hơn về VpTPD về mặt kỹ thuật chẩn đoán và cơ chế gây bệnh cũng như để phòng ngừa và kiểm soát TPD.
HÌNH ẢNH:
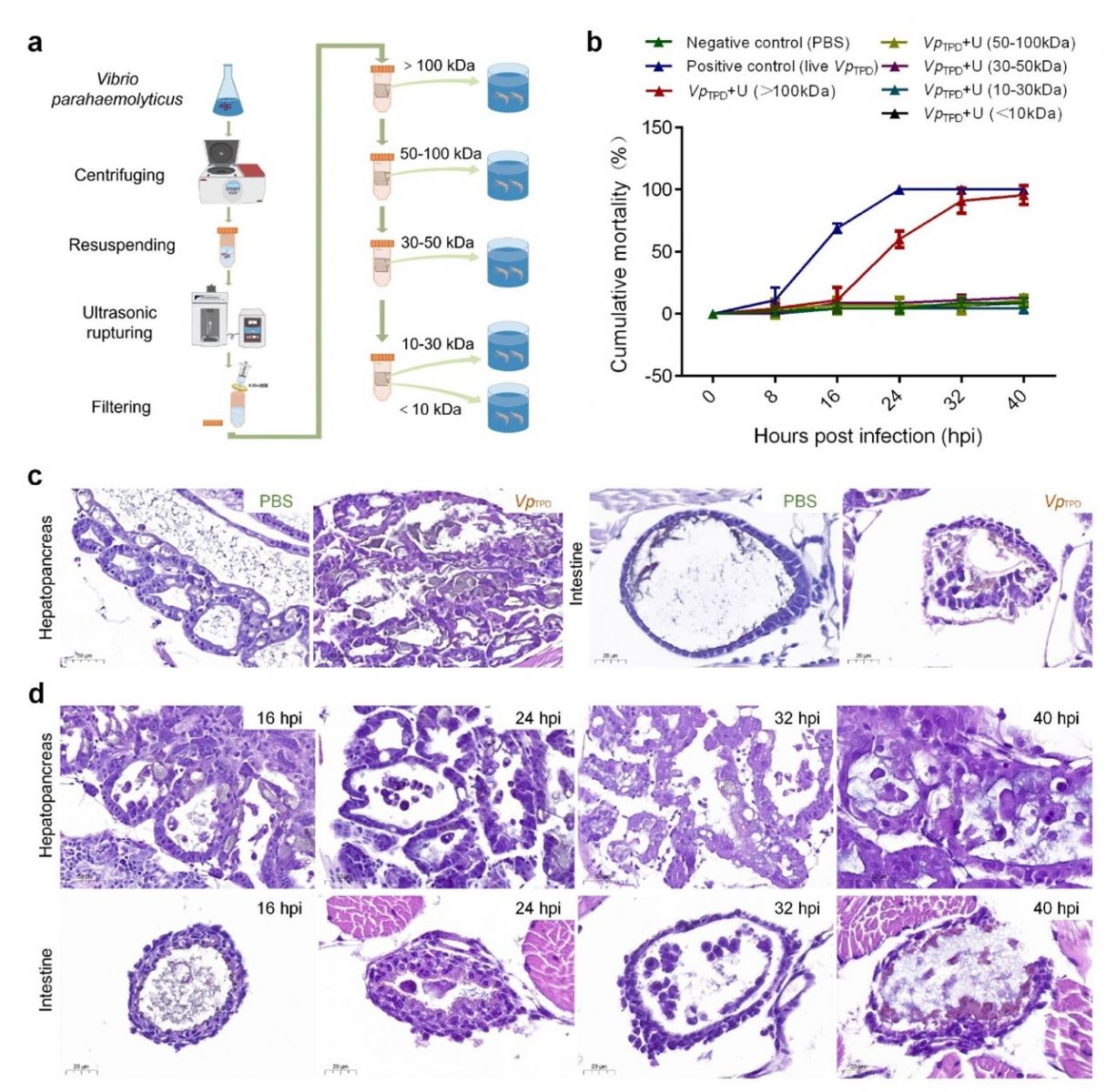
Hình 1 Phân tích khả năng gây bệnh của protein VpTPD có trọng lượng phân tử khác nhau đối với hậu ấu trùng Penaeus vannamei.
( a ) Sơ đồ các nghiệm thưcs được sử dụng để thu được protein VpTPD có trọng lượng phân tử khác nhau.
(b) Tỷ lệ tử vong tích lũy của hậu ấu trùng P. vannamei gây ra bởi các trọng lượng phân tử khác nhau của protein VpTPD trong thử nghiệm cảm nhiễm ngâm.
(c) Hình ảnh mô bệnh học của gan tụy và ruột của hậu ấu trùng P. vannamei từ nhóm cảm nhiễm VpTPD sống (đối chứng dương tính) và nhóm cảm nhiễm 1× PBS (đối chứng âm tính).
(d) Hình ảnh mô bệnh học của gan tụy và ruột của hậu ấu trùng P. vannamei từ nhóm VpTPD + U (>100 kDa) ở các thời điểm khác nhau sau khi nhiễm bệnh (bao gồm 8 hpi, 16 hpi, 24 hpi, 32 hpi và 40 hpi).
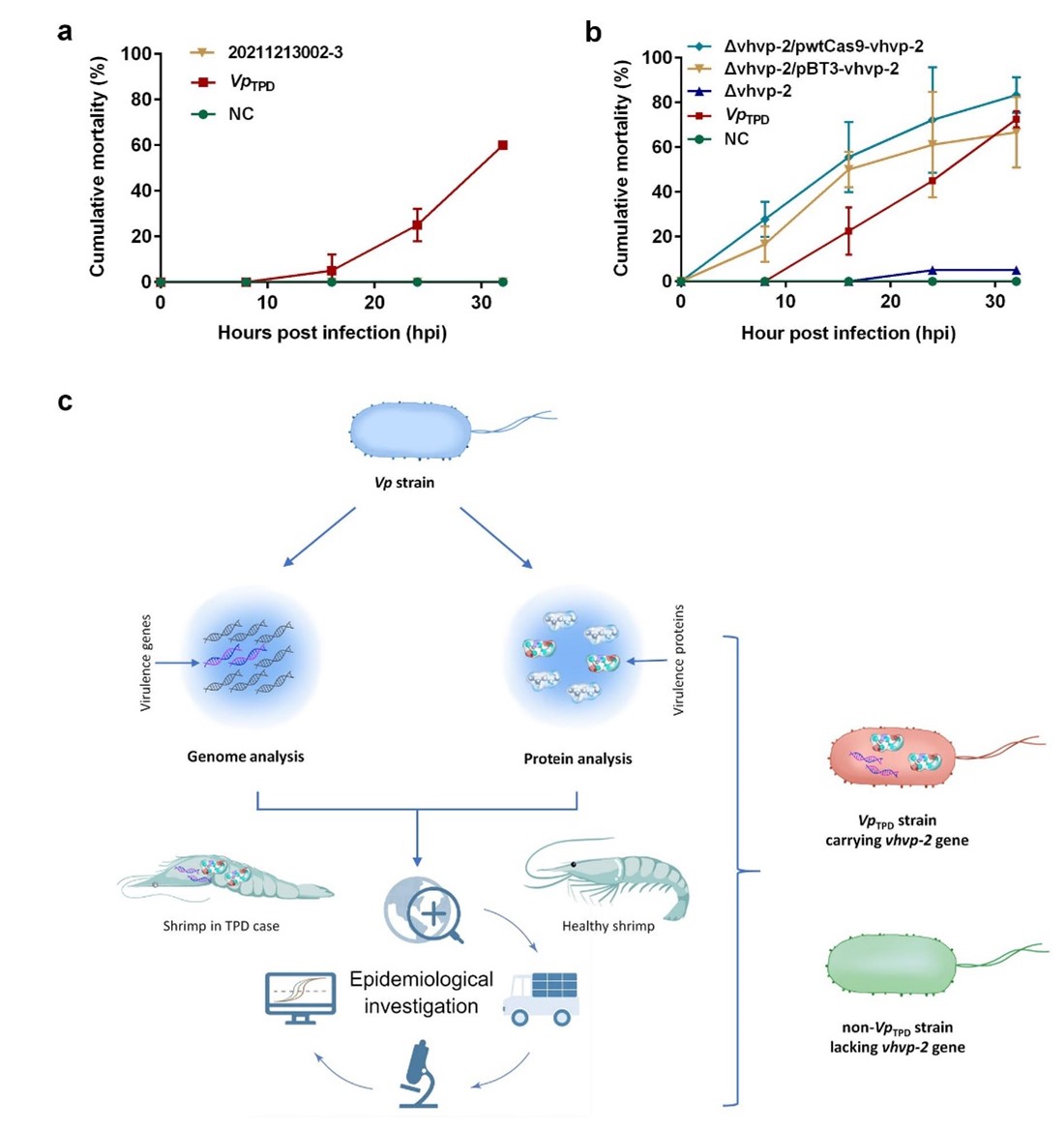
Hình 5 Xác định yếu tố độc lực chính của VpTPD
(a) Tỷ lệ tử vong tích lũy của hậu ấu trùng Penaeus vannamei ngâm trong chủng NC, VpTPD và Vibrio parahaemolyticus 20211213002-3. Hậu ấu trùng P. vannamei được ngâm với chủng Vibrio parahaemolyticus hoang dại VpTPD mang gen vhvp-2 hoặc chủng Vibrio parahaemolyticus 20211213002-3 thiếu gen vhvp-2 ở cùng liều lượng mầm bệnh. Tôm được theo dõi hàng ngày về tỷ lệ tử vong. Tỷ lệ tử vong tích lũy của tôm được biểu thị bằng giá trị trung bình và SD của hai dữ liệu lặp lại cho mỗi nhóm thử nghiệm.
(b) Tỷ lệ tử vong tích lũy của hậu ấu trùng P. vannamei ngâm ở các nghiệm thức
(c) Sơ đồ các quy trình được sử dụng để xác định yếu tố độc lực chính của VpTPD
Lược dịch: Thạc Sĩ . Trần Hữu Hậu - Viện Công nghệ Sinh học và Thực phẩm - Trường Đại Học Cần Thơ
Nguồn: Shuang Liu, Wei Wang, Tianchang Jia, Lusheng Xin, Ting-ting Xu, Chong Wang, Guosi Xie, Kun Luo, Jun Li, Jie Kong, Qingli Zhang., 2023, "Vibrio parahaemolyticus becomes lethal to post-larvae shrimp via acquiring novel virulence factors". Microbiology Spectrum - 10.1128/spectrum.00492-23.
- Mọi điều cần biết về EHP – Phần 1: Tổng quan về EHP
- Động lực lây truyền của virus hội chứng đốm trắng trên tôm thẻ chân trắng
- Các chủng Bacillus khác nhau được phân lập từ tôm sống sót sau AHPND có thể làm giảm tỷ lệ tử vong như thế nào?
- Nghiên cứu cho thấy tôm có thể đánh bại bệnh đốm trắng bằng cách di chuyển đến vùng nước ấm hơn.
- Mầm bệnh virus đốm trắng xâm nhập vào tôm qua cơ quan bài tiết nephrocomplex
- “Bọng nước” trên mang tôm
- Liên quan giữa nitrit và amoniac với tỷ lệ EHP trên tôm chân trắng và các vấn đề khác
- Liên quan giữa hội chứng phân trắng và EHP





